完了,這回我眞看出問題了。還是序列比對,之前有個地方我沒注意,就是S蛋白(spike)中閒有一個內切位置,在病毒進入細胞後,蛋白內切酶把S蛋白切成兩個亞基,在很多冠狀病毒一般是由胰蛋白酶(trypsin)完成的,包括SARS等。
這次問題就是在這個內切位置上了。之前我比對序列的時候,沒有留意這個位置。而這個位置正好是Covid-19和其它所有毒株,包括與其最近的雲南蝙蝠冠狀病毒 RaTG13都不同的(穿山甲在這段和Covid-19更遠),而且這前後的序列相對都比較保守(圖1,第1條序列是Covid-19,第2條是雲南蝙蝠RaTG13,第6條是人SARS)。Covid-19插入了4個氨基酸殘基(PRRA,即脯氨酸-精氨酸-精氨酸-丙氨酸),而前後的序列相比RaTG13都沒有變化。4個殘基就是12個核苷酸鹼基,並不是一種很容易發生的突變。而這一突變正好造成了一個furin內切酶識別位點。
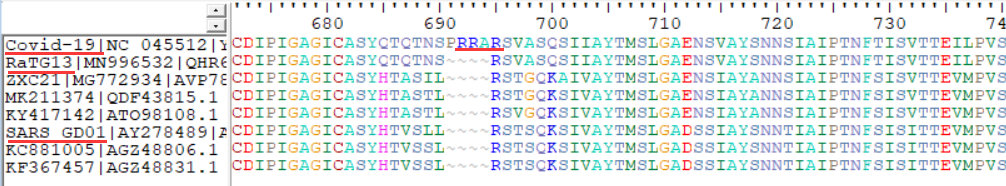
furin是一種蛋白內切酶,識別的蛋白序列爲RXXR(R是精氨酸,X是任一殘基)。PRRA加上後面緊跟着的R正好形成了furin識別位點。我覺得有問題的頭一點是在保守區引入了這樣一個插入,形成了識別位點。(之前我寫過一篇東西,說印度團隊的分析不靠譜,是因爲他們分析說有插入的地方太不保守,不同毒株什麼樣的長度都有。但這次是只有Covid-19與別的株不同。)因爲這個地方正好是切開兩個蛋白亞基,多兩個少兩個殘基一般並不很影響蛋白構象。
再看,這個位點會不會是被人有意引入的呢?結果發現,2009年眞就有人做過實驗了,發在PNAS上,就是把SARS在這個位置的RSTSQ變成了RRSRR,即引入了furin識別位點(原先是胰蛋白酶(trypsin)的識別位點)。結果發現病毒侵染效率提高了(圖2)。

所以,這個地方眞的很像人工設計的了。不過現在還沒有一個除這個位點以外其它部分都特別像Covid-19的母本病毒序列被公佈。
參攷文章:
- 李鑫 (2020) 武汉2019冠状病毒S蛋白可能存在Furin蛋白酶切位点
- Wong MC (2020) Evidence of recombination in coronaviruses implicating pangolin origins of nCoV-2019
- Belouzard S (2009) Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites
以上爲我發在新浪微雹的原文。
後面我補充的是:
- 這個多出來的PRRA確實不像自然演化的產物,而很符合人爲設計的特徵,但我也不能完全說這就不是自然演化,碰巧了。
- 本文我沒有指責任何人,病毒是無意或惡意放出並不知道,在武漢爆發也不說明就是中國人放出。
文章發出半天之後,我迫於一些不方便說的壓力,在微雹上隱藏了這篇文章。
這篇發出以後,我收到了微雹上另外幾位老師的回覆討論,包括 @內含子 的評論 和 @fengfeixue0219 的評論 。兼聽則明,我也推薦我的讀者能來看一下這兩篇,我認爲這是很有價值的討論,我也能從中學習到自己不熟悉的知識。我盡管未能被他們說服,但有一些點還是可以說一下的:
- 雖然我之前也寫過兩篇東西,說新冠病毒不像人爲製造的,那是當時我能看到的證據。一次是印度團隊提出新冠病毒的S蛋白前段的幾個插入被認爲是與HIV有關,還有一篇是James Lyons-Weiler提出新冠病毒是由不同病毒拼接而成,而且用了一個pShuttle-SN vector,我認爲這兩個東西的分析方法都有問題,見 《武漢冠狀病毒是人工改造插入了HIV片段?》和《武漢肺炎病毒爲人工製造可能性的探討》,所以他們提出的以我來看都不是有效的說明新冠病毒爲拼接而成的證據。而這次討論的PRRA的furin識別位點的證據和上面說的有所不同。
- 突變可以是人爲設計,也可以是自然演化。區別在於,自然演化的突變一般是在基因組全序列的各處隨機、散在地分佈,隨機突變完了,哪個能活下來繁衍後代,物競天擇適者生存之後你就能觀察到哪個。而如果是人設計一個東西,按照工程學的原理,應該希望產品功能可控、可預測、成功率高,這樣人盡量通過較少的操作來實現改造。因此如果說兩個序列同樣有n個位點的差異,如果這些位點很隨機地分佈在各處,就會更像是自然演化,而如果這些位點很密集分佈在某個區域,且功能以現有的方法能預測,就更像是人爲的。
- 兩位老師的回覆提到自然選擇也可能造成插入,這個我是同意的。所以我也只能說是像設計的,而不敢確定就是。
- 這種奇特的插入突變如果能較大幅度增強病毒的自我繁殖能力(受到正選擇),那麼即使這種插入突變發生的機率遠小於單鹼基替換,我們也是會看到這種突變的。
- 具體這種插入突變是小概率事件,以目前的瞭解可能很難計算其概率是多少。任何一個生物你讓它自然演化兩次,各產生幾個突變,都不會形成完全相同的序列。假如是不受到選擇的中性位點(比如外顯子的同義突變),某區域發生多少個突變的概率還可以算一下,而發生正選擇的區域會有蝴蝶效應,就沒法計算了,也就是說不能因爲是小概率事件而否認自然發生的可能。寄生生物,包括寄生蟲、致病菌、病毒等,爲了適應環境的變化,其演化速率非常不穩定,不適用分子鐘假設。
- 這是一個削弱新冠病毒爲人爲設計的理由:爲了能盡可能瞭解和掌握人工改造病毒的功能(不論是爲了硏究還是製造生物武器),人工改造會盡量選擇已經瞭解比較全面的株作爲母本,而不應該去隨便找個功能不明的野生株作爲母本,這樣風險太大,也難以衡量人工改造的效果。假如存在一個除furin識別位點以外其它部分都特別像Covid-19的母本病毒,其序列都沒公佈過,一定不可能是一個硏究很透徹的病毒。
- Covid-19病毒中,穿山甲病毒比雲南蝙蝠病毒RaTG-13更相似的片段爲S蛋白的RBD區域,而在插入的PRRA前後,Covid-19仍然是和RaTG-13更爲接近。下圖來自Wong MC (2020)。

如對本文有任何疑問,歡迎通過新浪微雹、telegram等渠道與我討論。

